
Abstract
Multi-cropping was vital for provisioning large population centers across ancient Eurasia. In Southwest Asia, multi-cropping, in which grain, fodder, or forage could be reliably cultivated during dry summer months, only became possible with the translocation of summer grains, like millet, from Africa and East Asia. Despite some textual sources suggesting millet cultivation as early as the third millennium BCE, the absence of robust archaeobotanical evidence for millet in semi-arid Mesopotamia (ancient Iraq) has led most archaeologists to conclude that millet was only grown in the region after the mid-first millennium BCE introduction of massive, state-sponsored irrigation systems. Here, we present the earliest micro-botanical evidence of the summer grain broomcorn millet (Panicum miliaceum) in Mesopotamia, identified using phytoliths in dung-rich sediments from Khani Masi, a mid-second millennium BCE site located in northern Iraq. Taphonomic factors associated with the region’s agro-pastoral systems have likely made millet challenging to recognize using conventional macrobotanical analyses, and millet may therefore have been more widespread and cultivated much earlier in Mesopotamia than is currently recognized. The evidence for pastoral-related multi-cropping in Bronze Age Mesopotamia provides an antecedent to first millennium BCE agricultural intensification and ties Mesopotamia into our rapidly evolving understanding of early Eurasian food globalization.
Similar content being viewed by others
Introduction
Multi-cropping, defined here as the seasonally sequential production of multiple crops on the same land in the same year, is an agricultural technique aimed at diversifying and intensifying economic and subsistence-based food, fodder, and forage production1,2,3. Similar to the contemporary world, this form of agricultural production was vital for provisioning large urban centers and financing imperial ambitions across ancient Eurasia4. For millennia, grain production in western Eurasia was limited to winter cereals, primarily wheat and barley, both of which were locally domesticated and adapted to Southwest Asia’s Mediterranean climate, with cool, wet winters and hot, dry summers. Summer grains and their wild relatives are not native to the region and thus summer cultivation only became possible with the translocation of millets and other East Asian and African domesticates.
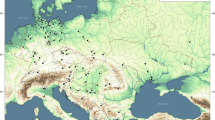
Millets represent a variety of fast-growing, small-seeded summer grains, initially domesticated in both Africa and northern China. Millets require summer rainfall (May–October > 120 mm) or irrigation5,6. Their short life cycle, drought tolerance, minimal maintenance, high returns, and protein-rich grains make millets versatile, nutritious, and labor-effective food sources for both people and livestock7,8,9. Two of the East Asian millets, broomcorn (Panicum miliaceum) and foxtail (Setaria italica) millet, were likely transported to western Eurasia both across the continent through Central Asia and along maritime trade routes (Fig. 1)5,10. Broomcorn (P. miliaceum), in particular, would eventually become one of the most important cereal grains in ancient Eurasia4.
By the mid-second millennium BCE, long-distance exchange networks connected all of Eurasia marking the near completion of the “Trans-Eurasian Exchange” in which East Asian domesticates arrive in Southwest Asia and Europe and wheat and barley reach East Asia11,12,13,14. Although domesticated millet is found throughout Central and South Asia and as far west as eastern Europe, cultivation of the crop is thought to be mainly restricted to areas with sufficient summer precipitation (Fig. 1)5. Even today broomcorn millet (Panicum miliaceum) is a very minor cultivar in Iraq15,16 and rarely grows ferally even in perennially damp places17. As a result, most archaeologists believe millet was only introduced to Mesopotamia (ancient Iraq) and other areas that lack summer precipitation with the construction of massive, state-sponsored irrigation systems during the mid-first millennium BCE, which would have made multi-cropping possible and worthwhile5,18. In contrast, textual evidence suggests millet may have been cultivated in Mesopotamia as early as the third millennium BCE, possibly being introduced via maritime routes from the Indus Valley10,19 or overland via expanding Bronze Age trade networks20,21,22.

Map of archaeological sites with archaeobotanical evidence for broomcorn millet (Panicum miliaceum) from the 3rd–1st millennium BCE. See Supplementary Table S2 for site data sources. Summer precipitation (May–October)23 is displayed in grayscale (after5). Red lines and arrows indicate domestication areas and translocation routes (after13,19), and black lines indicate later silk road corridors (after24). This figure was generated in Esri’s ArcGIS 10.6.1 (http://www.esri.com/software/arcgis) using Esri World Imagery (Sources: Esri, DigitalGlobe, GeoEye, i-cubed, USDA FSA, USGS, AEX, Getmapping, Aerogrid, IGN, IGP, swisstopo, and the GIS User Community).
Previous evidence for millet cultivation in Mesopotamia
Textual evidence
Mesopotamian textual specialists have long argued that millet was an important crop in the region as early as the third millennium BCE25,26. The Akkadian term for millet (duḫnu/tuḫnu) is first explicitly mentioned in mid-second millennium BCE cuneiform texts, found at the Mesopotamian sites of Nippur and Nuzi27. Texts from Nuzi suggest that millet was planted in conjunction with sesame: “plant sesame and millet! there is one homer of sesame and millet which is already planted,” or gifted as grain along with barley: “PN [personal name] gave to PN2, PN3, and PN4, four homers of barley and two homers of millet as…for the properties27.” Some scholars link duḫnu/tuḫnu to the Akkadian word arsikku (Sumerian: ar zig), possibly pushing references to millet back into the third millennium BCE26,28. Old Akkadian (third millennium BCE) texts also refer generically to both “early grain” (Sumerian: še nim; Akkadian: harpu) and “late grain” (Sumerian: še sig; Akkadian: uppulu), which was sown in spring and harvested in late summer and thus may refer to either millet or sesame26,29,30. These are the only known summer-sown “grains,” and certainly all the technology needed for their cultivation would have been available long before the second millennium BCE31.
Archaeobotanical evidence
Textual evidence contrasts sharply with archaeological perspectives that largely ignore the possibility or implications of millet multi-cropping in Mesopotamia prior to the first millennium BCE. Some archaeologists do not doubt that millet was also present in semi-arid Mesopotamia during the second millennium BCE because it is contemporaneously present in adjacent areas with more temperate climates32, and sesame, also a summer crop, is a known cultivar in the region from the third millennium BCE onwards6,33,34. However, millet is rarely mentioned in discussions of early Mesopotamian agriculture34,35 and is generally excluded from models of Bronze Age Mesopotamian food production36,37.
The contradiction between textual and archaeological perspectives is due to the nearly complete absence of archaeobotanical evidence for millet in Iraq in all periods38 and, until recently, its general scarcity in adjacent regions. The earliest unequivocal archaeobotanical evidence for broomcorn millet in Mesopotamia is from c. 700 BCE when millet grains are found in large numbers at Nimrud and Fort Shalmenesar (Fig. 1, no. 26; Supplementary Table S2)39. Citing Boserup40, Miller et al.5 argue that millet may have been known in Mesopotamia, but was absent due to ecological constraints (i.e., the lack of summer precipitation) and it was never used prior to the large-scale Neo-Assyrian imperial intensification systems, even as a diversification or risk reduction strategy (see also41). Likewise, Rosenzweig18 credits the Neo-Assyrians and their agricultural maximization policies with the introduction of millet and other non-local crops to northern Mesopotamia.
However, archaeobotanical remains are not entirely absent prior to the first millennium BCE. Earlier evidence for Panicum miliaceum may be present in impressions of millet grains on ceramics from the site of Jemdet Nasr (ancient Kish) in southern Iraq dating to 3000 BCE42,43, but the interpretation of botanical impressions has been argued to be unreliable44. A few charred Panicum grains were reportedly found inside a small jar from the same site45, and a single grain of P. miliaceum was identified from a secure Late Bronze Age (14th–13th cent. BCE) oven context at Gurga Chiya in the Shahrizor plain in northeastern Iraq46. In northern Syria, isolated grains of P. miliaceum are also reported in second millennium BCE contexts at Tell Sheikh Hamad47,48 and Tell Mozan in northeast Syria49; although the Tell Mozan finds are not mentioned in the final report50. The rarity of millets in archaeobotanical data from Bronze Age Mesopotamia has led archaeologists to interpret these finds as exotic imports, intrusive grains, or very minor cultivars, and thus millet has played almost no role in our interpretations of agro-pastoral production in the region.
Second millennium BCE Khani Masi
This study presents new data from the site of Khani Masi, located along the Upper Diyala/Sirwan River, a tributary of the Tigris River, in the Kurdistan Region of Iraq (Fig. 2a). Khani Masi is composed of more than a dozen mounds clustered along a relict levee above the Diyala/Sirwan River. From 2014 to 2019, the Sirwan Regional Project (SRP) initiated a program of archaeological investigations focusing on large-scale excavation of a sprawling low mound (SRP 46), which measures c. 5 ha in area and was occupied exclusively during the mid- to late-second millennium BCE51,52,53,54. At this time, Khani Masi appears to have close cultural and economic ties to Kassite Babylonia, centered in southern Mesopotamia. Excavations have revealed a sequence of major construction episodes, with the earliest phases dated to around 1450 BCE and subsequent building phases during the fourteenth and thirteenth centuries BCE. Settlement at SRP 46 ended following the abandonment of an extensive baked mudbrick building complex around 1100 BCE, and there is no evidence that this part of the site was ever reoccupied51.
The region in which Khani Masi is located has a typical Mesopotamian steppe climate (Irano-Turanian vegetation), with cool, wet winters and hot, dry summers55, and regional paleoclimate data suggest that a similar climate system prevailed during the second millennium BCE (Fig. 2b)56. Today, average winter rainfall in the Khani Masi area is marginally sufficient for dry-farmed wheat and barley cultivation (Nov–April 1970–2000: 314 ± 51 mm23; 334.6 ± 115.3 mm57) (Fig. 2c). The high interannual variability in precipitation means that today, and probably during historic periods, irrigation was necessary to support reliable agriculture, even for the Southwest Asian crops, wheat and barley. From May through October, Khani Masi only receives an average of 17 ± 9 mm (1970–2000)23 and thus any summer cultivation would unquestionably require irrigation. Although direct evidence for irrigation works predating the first millennium CE have not yet been observed in the region, the area surrounding Khani Masi (a Kurdish name meaning “spring of the fishes”), is replete with perennial, spring-fed streams, supplied by groundwater originating in the Zagros Mountains to the northeast. This well-watered plain has a rich history of human occupation dating back to the Neolithic period58, and thus it is reasonable to conclude that irrigation was likely practiced for many millennia.

The Khani Masi region and its environmental context. (a) Map of Upper Diyala/Sirwan River region with perennial water sources, growing season rainfall isohyets (November–April)23, agricultural zones59, and climate proxy sources indicated. This figure was generated in Esri’s ArcGIS 10.6.1 (http://www.esri.com/software/arcgis) using Landsat 8 imagery (4 Oct 2021; courtesy USGS and NASA). (b) Paleoclimate speleothem record of the last c. 4000 years from the Kuna Ba cave (Kurdistan Region, Iraq; a)56 highlighting the Kassite (c. 1550–1150 BCE) and Neo-Assyrian periods (c. 10th–early seventh centuries BCE). (c) Averaged monthly Diyala/Sirwan River discharge volume from meter stations north (T16; a) and south (T17; Fig. 2A) of Khani Masi for the years 1931–1955, prior to the Darbandikhan and Hamrin dam constructions60. The black line indicates average (± SD) precipitation by month from 1960 to 201657 compared with the agricultural cycles of major crop types by month for Iraq16.
In 2019, the SRP excavations near the center of the site uncovered what appear to be a large, deep midden deposit (Trench Y82, Fig. 3). Excavations of midden deposits are not uncommon in the greater ancient Near East (e.g.61,62), but have rarely been studied in detail in Mesopotamia54,63,64. Abundant ceramics and AMS carbon-14 samples securely date the deposit to the mid-late second millennium BCE (Figs. 3, 4 and Supplementary Tables S3–4). During phytolith morphological analysis of the sediments from Trench Y8254 (see Supplementary Text), we encountered phytoliths in anatomical connection, or multicellular structures (also known as silica skeletons66 or articulated groups67), composed of interdigitating phytolith morphotypes68,69 similar to Panicum miliaceum in ten samples (Table S1). In general, phytolith preservation was good across all samples (Supplementary Text) as is typical for archaeological sites (or tells) in Southwest Asia where phytoliths are removed from the silica cycle70,71,72,73. No interdigitating phytolith morphotypes distinctive of domesticated Panicum sp. were observed in surface control samples, and millet is not currently cultivated in the area. Thus, modern contamination should be excluded.

Eastern profile of Khani Masi Trench Y82. Colors represent major stratigraphic phases, or depositional episodes. Circles indicate sediment sample locations and approximate locations of excavated charcoal samples (Supplementary Tables S1 and S3). Samples with measurable interdigitating phytoliths in anatomical connection (silica skeletons) are highlighted in red. Figure modified from54. This figure was generated using Agisoft’s Metashape Professional Software v. 1.5.3 (http://www.agisoft.com/).

OxCal65 multiplot of Bayesian modelled 14C dates by phase (indices: Amodel 100.1, Aoverall 100.9). The modeled start date for phase 4, stratigraphically earlier than sample 59, is 1571–1322 BCE (± 2σ, 95.4% confidence) (Supplementary Tables S3–S4). Asterisk denotes new dates for this study. Unmodelled dates were previously reported by Laugier et al.54.
Independent of the analyses performed by Laugier et al.54 (supplementary text, Table S1, and Figure S1), in this study, we performed a morphometric analysis on 30 multicellular structures composed exclusively of interdigitating phytoliths from four samples (Fig. 3) to determine their taxonomic origin (“Material and Methods” section). interdigitating phytoliths in anatomical connection were abundant and easily photographed in sample 59 (N = 27), while samples 67, 73, and 75 yielded only a single interdigitating multicellular structure each. Providing context to these four samples, samples 59, 73, and 75 are from burned animal dung-rich deposits while sample 67 is from an outdoor surface (Supplementary Text and Table S1). Sample 59 is the earliest dung-rich sediment in the Trench Y82 sequence, and post-depositional organic decay is indicated by the presence of authigenic phosphate54. Phytolith morphotype results showed that riparian vegetation like sedges (Cyperaceae) were rare (< 2.1%) (Supplementary Text and Fig. S1). bilobate short cells, distinctive of Panicoideae (C4) vegetation like millet, represent 30% of the short cell assemblage in sample 59 (the most of any sample at Khani Masi) and between 9.3 and 10.4% in samples 67, 73, and 75 (Supplementary Text and Fig. S1)54.
Although some scholars argue that morphometric analysis is not necessary to securely differentiate Panicum miliaceum inflorescence bracts (upper lemma and palea) from other domesticated millets74, P. miliaceum and other domesticated millets share several features with their wild relatives and other weedy Panicoideae (C4) grasses75,76. For example, the Panicum species, Panicum bisulcatum (Japanese panicgrass) and Panicum repens (torpedo grass), are morphometrically very similar to P. miliaceum and can only be distinguished from broomcorn millet based on a single criterion77,78. P. bisulcatum is not native to Iraq, but P. repens is native and present in riparian areas17,79,80. In fact, five of the nine genera representing millets and their wild relatives81 are present in modern Iraq (Digitaria, Echinochloa, Panicum, Paspalum, Setaria, and Sorghum) (Supplementary Table S6)17,80. Thus, it is necessary to use at least five diagnostic criteria to ensure secure species-level identifications of millet inflorescence phytoliths (Supplementary Table S5)68,75,76,77,82. Like phytoliths from grass inflorescences, bilobate short cells can be used to distinguish between domesticated millet species83. However, because the primary concern in this study was differentiating P. miliaceum from wild panicoids, different bilobate short cell types were not analyzed here.
Using five criteria, Panicum miliaceum inflorescence phytoliths can be confidently differentiated from all other known millet-like panicoideae species native to Iraq (Supplementary Fig. S2 and Table S5–6)68,76,77,78,82,83,84,85,86. That is, based on the current knowledge of phytolith morphometrics and the native ranges of species that produce interdigitating phytolith morphotypes, P. miliaceum phytoliths are distinct from all other known phytolith reference species except Panicum miliaceum L. subsp. Ruderale (Kitag.) Tzvelev, the debated progenitor, feral relative, or weedy companion of domesticated P. miliaceum78. Panicum ruderale is not native to Iraq and thus its presence would also indicate human translocation.
Results
This study identified 30 phytolith multicellular structures (silica skeletons), with characteristics consistent with the inflorescence bracts of broomcorn millet (Panicum miliaceum) (Figs. 5, 6 and Supplementary Table S7). Multicellular structures sizes range between 1 and 10 individual measurable interdigitating phytolith morphotypes (average size: 3) for a total of 90 individually measured phytoliths. Note that partial or broken morphotypes at the edges of the multicellular structures were not measured. 28 of the 30 interdigitating multicellular structures meet five criteria and 19 meet all six criteria (Supplementary Table S7). In every interdigitating multicellular structure, papillate phytoliths are absent (criteria 1, Supplementary Table S5), margin processes are η-type (criteria 2), and the process height to body ratio is greater than 1 (criteria 6). 13 multicellular structures have distinctly finger-type endings, and 14 have either both ending types or intermediate appearances (criteria 3). The overall averages for both long cell (elongate) ending lengths (W = 8.13 ± 1.10 µm, criteria 4) and the ratio of process height to ending length (R = 0.71 ± 0.06 µm, criteria 5) are within one standard deviation of the values reported by Lu et al.77 for P. miliaceum. All measurements of both the individual and multicellular interdigitating phytoliths exceed the minimum required sample size for ensuring means are within 5% of actual population means at a 90% confidence level (Supplementary Table S8)87,88.

Microscope images of a selection of measured interdigitating multicellular phytoliths. (a–e) interdigitating multicellular structures from sample 59 that contain long cells (elongate) with ‘finger-type’ endings and η-type levels II–III margin undulation patterns. (f) interdigitating multicellular structures from sample 59 with shorter ‘wave-type’ endings and η-type levels II–III margin undulation patterns. Scale bars are 50 µm.

A plot of average morphometric values, R and W, for each Khani Masi interdigitating multicellular structure (red and yellow) compared to values reported by Lu et al.77 (light gray circles and triangles). W (x-axis) is the long cell (elongate) ending length (Table S6, criteria 4), and R (y-axis) is the ratio of processes amplitude to endings (Table S6, criteria 5). The distribution of Khani Masi multicells with exclusively ‘finger-type’ long cell endings (yellow) falls completely with the normal distribution ellipse for broomcorm millet as reported by Lu et al.77 (dark gray). All points represent average values with standard deviation error bars. Khani Masi (red and yellow) point size indicates the level (I–III) of the η-type undulation pattern. Normal distribution ellipses are colored by broomcorn millet (dark gray), foxtail millet (blue), all Khani Masi multicells (red), and only Khani Masi multicells with ‘finger-type’ long cell endings (yellow). Figure modified from Lu et al.77.
Of the species native to Iraq with known interdigitating inflorescence phytoliths, only Paniucm repens (torpedo grass) shares four of the same diagnostic criteria with Panicum miliaceum. P. repens only produces short, ‘wavy-type’ endings, whereas P. miliaceum mostly produces ‘finger-type’ endings (criteria 3)77,78. Nineteen interdigitating multicellular structures have finger or both finger- and wavy-type endings making them potentially distinct from P. repens. However, cautiously using the stricter criteria of exclusively ‘finger-type’ endings, eleven multicellular structures are securely identified as P. miliaceum (highlighted yellow in Fig. 6). Because several species can produce η-type level I–II margin processes, an even more conservative identification of P. miliaceum could also require η-type level III margin processes. In this case, six interdigitating multicellular structures have both η-type level II–III or III margin processes and exclusively ‘finger-type’ endings, increasing confidence for the identification of P. miliaceum at Khani Masi.
It is possible that the interdigitating phytolith morphotypes analyzed in this study could potentially be produced from some yet unknown wild C4 plant that has phytoliths identical to those of Panicum miliaceum. However, based on the current phytolith knowledge, the interdigitating phytolith morphotypes measures fit remarkably well within the broomcorn population. For example, Panicum turgidum Forssk. (desert grass) is not well studied from a phytolith perspective, although it does not appear to resemble the strict criteria for P. miliaceum89,90. Panicum turgidum is also adapted to and moderately present only in the sandy desert regions in the extreme southeast of Iraq (Saharo-Sindian), and is not expected to grow in the moister Khani Masi region17,91. Future research should focus on generating accessible phytolith references for the wild grasses of Iraq and the greater Mesopotamian region.
Discussion
Our results demonstrate that broomcorn millet (Panicum miliaceum) is present at the mid-second millennium BCE site Khani Masi located along the Upper Diyala/Sirwan River in northern Iraq. This result represents the first phytolith identification of P. miliaceum from ancient Iraq and aligns with both contemporary textual sources and some regional macrobotanical evidence that suggests millet was present in Mesopotamia at this time.
The presence of millet at Khani Masi may also provide the earliest evidence to-date for regional cereal multi-cropping in Mesopotamia. That is, in addition to the winter cereals, wheat and barley, which are attested in both the micro- and macro-botanical records at the site51,54, we also now have robust micro-botanical evidence for summer grains. Although the presence of a plant is not enough to prove it was cultivated92, the discovery of a non-native, east Asian domesticate outside its environmental niche and within a dung-rich context strongly suggests that it may have been cultivated as a forage crop. If cultivated locally at Khani Masi, millet-as-forage provides a “long prelude”93 or “clear antecedent”41 for its use in the region’s first millennium BCE agricultural intensification systems. Locally cultivated millet also suggests that previous arguments for environmental constraints on early millet cultivation may be overstated and require alternative explanations.
While Mesopotamia is (semi-)arid in terms of precipitation, it contains an abundance of perennial water sources that could support summer cereal cultivation without major investments in irrigation. As further articulated below, our results suggest that the absence of evidence for millet in previous investigations is due to the particularly strong taphonomic bias against millet grain preservation as well as to the low archaeological visibility of pastoral lifeways. Thus, millet cultivation was likely far more widespread in the second millennium BCE than is currently recognized. Furthermore, the availability of millet as an alternative food source for people or animals requires the reassessment of Bronze Age models of urban provisioning, resilience, and human-environment interactions.
Summer cultivation in (semi-)arid Mesopotamia
Mesopotamia (ancient Iraq) has a long history of irrigation and a myriad of perennial water sources, particularly in the ecologically diverse Zagros foothills zone, and the cultivation of millet in the region should be unsurprising. Irrigation technology was developed in the region during the sixth–fifth millennium BCE94,95 meaning it had been practiced for millennia by the second millennium BCE. That sesame is cultivated in the region from the third millennium BCE onwards further emphasizes that Mesopotamian communities were familiar with summer crops and were versed in small-scale summer cultivation. Thus, while the local seasonal climate may be unfavorable for precipitation-based summer cultivation, river discharge rates during April–June suggest that preexisting irrigation infrastructure could easily have been used to irrigate millet sown as a cover crop in river flood control areas or in fallow fields16,60,96 (see Fig. 2c). Summer crops also could be grown opportunistically outside winter cultivation areas along perennial water sources such as riverbanks and karstic springs.
Two alternative scenarios, although unlikely, could explain the consumption of millet by animals at Khani Masi without local cultivation: (a) the feral growth of the domesticated grain near perennial water sources or (b) the long-distance transport of unhulled grains from their natural growing range.
First, the feral growth of domesticated Panicum miliaceum around nearby perennial water sources is unlikely but cannot be completely ruled out. Feral growth requires that broomcorn millet was introduced into the region before the midden was formed at Khani Masi, during late third–early second millennium BCE. If this was the case, the implication is that domesticated Broomcorn millet was still known and actively used for animal forage at this time, centuries earlier than previously accepted.
Second, Khani Masi is located along a strategic trade route connecting lowland Mesopotamia to the Iranian Zagros highlands and Central Asia51,52. Yet, it is unlikely that sufficient amounts of unhulled millet grains would have been transported hundreds of kilometers from the Taurus-Zagros mountains or from maritime ports in lowland Mesopotamia only to be used as fodder for local sheep-goats (Supplementary Fig. S3). Moreover, the maximum estimated one-way travel distance for sheep and goat herds based on average consumption to defecation times is 35–47 kilometers97 and does not allow for a scenario in which millet was consumed in its natural range and deposited at Khani Masi (Supplementary Fig. S3). Thus, the most parsimonious explanation is that a small number of seeds were transported and planted locally. Furthermore, the presence of Panicum miliaceum in multiple layers in Trench Y82 suggests it was cultivated in small quantities over multiple years (Table S1).
Factors affecting broomcorn millet preservation and recovery
By the mid-second millennium BCE, Mesopotamia is demonstrably integrated into the globalized networks that connected all of Eurasia98,99,100. The technology and ecological niches required for summer cultivation were present, and both millet and sesame are mentioned concurrently in textual sources. Macrobotanical evidence of millet has been scarce across Mesopotamia, but now millet micro-remains are verified at Khani Masi. Together, these lines of evidence suggest that a combination of taphonomic and cultural factors are affecting the regional recovery of millet.
Taphonomic factors
As with most archaeological material, several factors condition both the entrance of plant remains into settlement areas and their preservation after arrival101. In other ethnographic and archaeological case studies, the lack of macrobotanical evidence for millet cultivation has been attributed to its minor cultivation, processing in off-site areas, and particular susceptibility to destructive taphonomic processes92,102. While millet may have been a minor crop in second millennium BCE Mesopotamia, its paucity in the long-term archaeobotanical record is most likely the result of the fragility of millet grains coupled with regionally poor macrobotanical preservation.
Compared to other crops, millet’s small inflorescence structures and high fat, oil-rich grains make it particularly susceptible to destruction during charring5,92,103––the primary mechanism for chaff and grain preservation in most regions104,105. Accordingly, multi-proxy methods are required to investigate millet processing even in regions where millet is a major cereal crop102,103. Notably, in regions where millet was introduced and not anticipated, highly degraded charred grains may be mistaken for weeds44.
In Southwest Asia, preservation conditions for macrobotanicals can be poor, especially in shallower sites, and the ubiquities of even the major crops, wheat and barley, can sometimes be too low for meaningful statistical analysis (e.g.106). For millet, macrobotanical finds are remarkably rare in all periods, even in periods when millet is intensively cultivated38. Millet finds from Southwest Asia seem to be restricted to a single grain for an entire site (e.g.46,49,107) or large caches recovered under exceptional preservation conditions such as roof storage collapse from catastrophic fires (Tille Höyük, Turkey and Haftavan, Iran32); or in jars with tar (Nimrud39).
In many ways, the archaeobotanical record for millet mirrors that of sesame (Sesamum indicum L.), another small-grained, oil-rich summer plant translocated into Mesopotamia. Like millet, sesame seeds are also textually attested but exceedingly rare in the archaeobotanical record29. Their small size and high oil content make carbonized sesame seeds extremely fragile and prone to disintegrate during the recovery process. Further, their relatively small quantities and processing in off-site areas make them less likely to enter the archaeological record in the first place29,108. Consequently, no sesame grains are attested in Mesopotamia for the nearly 1000 years between the earliest grains recovered ca. 2300 BCE (Tell Abu Salabikh, Iraq) and those dating to the late second millennium BCE6,33,109. Where data are published, sesame finds, too, are restricted to very few grains or large caches29. However, like millet, recent proteomic, residue, and microbotanical approaches are demonstrating that sesame and other exotic plants were more widespread in second millennium BCE Southwest Asia than previously thought100,110,111.
Cultural factors
The context in which millet was recovered at Khani Masi suggests an additional taphonomic reason why millet is rare in the second millennium BCE: its primary use as animal forage (or fodder). In contrast to sesame, millet phytoliths at Khani Masi were primarily recovered from burned and discarded dung-rich sediment that suggests introduction via animal dung54. Archaeologists and biologists alike have long appreciated the facts hidden in animal waste112,113, but the value of dung and its contents is still underappreciated for investigating Mesopotamia’s economies and ecologies.
Dung-associated plant material is subjected to additional destructive processes that decrease the likelihood of grain identification from macrobotanicals. First, dung is most likely to enter the archaeological record through fuel use and animal penning (although evidence is still pending for dung as a common construction material in Mesopotamia). Second, unlike wild seeds, which are abundant in ruminant animal dung, domesticated cereal grains are starchy or oil-rich with thin protective outer coatings and rarely survive sheep and goat digestion114,115,116. Third, because dung in ancient settlements is often used as fuel or burned to reduce the volume of dung accumulating in animal penning areas, any fragile millet grains that survive digestion would be subsequently destroyed through burning. Finally, discarded organic rich dung and ashes often decay after deposition (diagenesis) further destroying organic macrobotanical evidence114.
The strong taphonomic bias against millet grain preservation means that this grain has been below our ability to resolve using traditional macrobotanical methods77. Microbotanical and geochemical approaches (e.g., phytoliths, dung spherulites, FTIR), which can effectively identify animal dung and its contents, have not yet been widely used in the region. This study demonstrates, however, that millet was cultivated in Mesopotamia and that phytolith analyses of dung deposits are likely key for investigating the role of forage and fodder in the advent of regional multi-cropping.
The pastoral origins of multi-cropping in Mesopotamia
The recovery of millet from animal dung—consumed as a forage crop—suggests that the initial practice of multi-cropping in Mesopotamia is likely associated with small-scale pastoral diversification strategies—not imperial agricultural mandates. Pastoral, here, is defined broadly as the husbandry of sheep-goats (after117,118), acknowledging that local pastoral systems and their specific practices, level of mobility, and integration into agricultural systems vary widely. Millet grown as a forage crop would be directly consumed by animals, not harvested. The pastoral origins of multi-cropping in Mesopotamia complement multiple botanical and isotopic studies from across Central and South Asia that also suggest millet was adopted slowly, through bottom-up, pastoral initiatives13,22,41,93,119,120,121,122,123. Millet’s low investment, high return qualities made it especially well-suited to the needs of the semi-mobile pastoralists who transported it across Central Asia’s ecologically diverse landscapes5,124. It is fitting then that this new crop may have been first adopted by pastoralists living in the environmentally complex Mesopotamian-Zagros interface.
Pastoral practice outside of institutional spheres has been a topic of intense debate in Mesopotamian archaeology because it is not well documented in Mesopotamia’s archaeological or textual records125. However, we should consider that the introduction of new foods and related practices likely disrupted lifeways18. As well as enhancing agro-pastoral resilience through diversification, millet may have been a destabilizing force by offering increased autonomy from established (or distant) socio-political and economic systems126,127. In both cases, the possible pastoral origins of multi-cropping highlight the influence of steppe region pastoral practice on the political and land use histories of Southwest Asia. Many Southwest Asian crops and animals were first domesticated in the Zagros foothills (“hilly flanks”)128,129, and this study suggests that the Zagros foothills may have continued to be a regional center of agro-pastoral innovation for Mesopotamia during the Bronze Age.
Reassessing provisioning models in light of food globalization
Beyond the Zagros Region, the adoption of millet likely had far-reaching impacts on Mesopotamia’s social, political, and economic systems beginning in the second millennium BCE. The verified presence of millet in mid-second millennium BCE Mesopotamia sheds new light on historical events and trajectories of the region and requires a reassessment of models of urban provisioning, resilience, and human–environment interactions. For example, Lawrence et al.36 attribute the “decoupling” of urban site size (and population) with climate trends after 2000 BCE and urban size with sustaining area after 1200 BCE to changes in labor organization, taxation, and integration into long-distance trade networks. However, like most models of Mesopotamian economies, they have not yet explicitly considered the impact of new crops36,37. However, this decoupling of demographic and environmental variables coincides with the arrival of new crops with properties optimally suited to diversifying and strengthening the resilience of Mesopotamia’s agro-pastoral production systems. Even a low-level or opportunistic cultivation of millet, for human or animal consumption, may have had a significant impact on urban provisioning and thus resilience capacity130. Future studies could further investigate the origins of multi-cropping by investigating isotopic δ13C enrichment from low-level millet (C4) consumption and by deploying microscopic methods that acknowledge the taphonomic biases against millet grain preservation. Perhaps uncoincidentally, evidence of millet cultivation is nearly as rare as studies using isotopic131 and phytolith approaches54 with potentially critical impacts on our understanding of Mesopotamia’s social, political, and economic systems.
Conclusion
Here we provide the earliest microbotanical evidence of broomcorn millet (Panicum miliaceum) in Mesopotamia (ancient Iraq) and suggest that the origins of multi-cropping (summer cultivation) begin in the second millennium BCE. This finding aligns with ongoing investigations of early food globalization across Eurasia, a conversation in which Mesopotamia has been notably absent. As in other regions, the initial use of millet in Mesopotamia was likely as a foraging crop. Agro-pastoralists in the Zagros-Mesopotamian interface may have grown millet opportunistically at low levels for centuries as a diversification strategy41,132 before it was considered food suitable for human consumption or economically advantageous12,93 to the political economies within the first millennium BCE Neo-Assyrian Empire. Strong taphonomic bias against millet grain preservation provides an explanation for why its recovery has been so rare despite its known presence in textual sources. Micro-remain analysis offers a promising path forward for exploring the processes and practice of multi-cropping in Mesopotamia. In fact, this study highlights that micro-remain analyses have the potential to fundamentally transform our understanding of daily life, the formation of states and empires, and human-environment relationships in one of the most prominent and strategic nodes of ancient Eurasian and African networks.
Materials and methods
Excavation and sampling
Two adjacent trenches (10 × 2.5m2) separated by a 0.5 m baulk were excavated in area Y82 at Khani Masi (SRP46) by the Sirwan Regional Project (SRP). Charcoal samples for 14C dating were collected during excavation and analyzed at the University of Arizona AMS Laboratory. 14C date ranges were calibrated using OxCal v4.4.4 (Fig. 4 and Supplementary Tables S3–4)65. Bulk sediment samples (~ 30 g) were collected in plastic bags directly from the freshly cleaned baulk section, and sampling tools were cleaned with acetone between every sample. Sample locations were tagged, photographed, and geolocated using an Emlid RS + RTK GNSS system.
Microscopy
Phytoliths were extracted using the Katz et al.133 method. Phytoliths were identified and photographed using a Nikon eclipse LV100N POL petrographic microscope at 200× and 400× magnification. Morphological identification followed the standard literature66,134,135,136,137 using the International Code for Phytolith Nomenclature (ICPN 2.0) when possible67.
Morphometric analysis
Quantitative phytolith measurements were taken in ImageJ software (version 1.5.3) using the morphometric criteria defined by76,77,82 (Supplementary Fig. S2 and Table S6). To avoid any taphonomical bias in the morphometric analysis, we measured only complete individual phytoliths forming multicellular structures (silica skeletons). Partial or broken individual phytoliths at the edges of each silica skeleton were not measured. Following Ball et al.87,88, minimum sample sizes were calculated for all measurements for both multicellular structures and individual phytoliths to ensure sample means were within 5% of the actual population means at a 90% confidence level (Supplementary Table S8).
References
Andrews, D. J. & Kassam, A. H. The importance of multiple cropping in increasing world food supplies. In Multiple Cropping (eds Papendick, R. I. et al.) 1–10 (American Society of Agronomy, 1976).
Gallaher, R. N. Multiple cropping systems. In Management of Agricultural, Forestry, and Fisheries Enterprises Vol. 1 (ed. Hudson, R. J.) 254–265 (Eolss Publishers Co., Ltd., 2009).
Petrie, C. A. & Bates, J. ‘Multi-cropping’, intercropping and adaptation to variable environments in indus south asia. J. World Prehistory 30, 81–130 (2017).
Spengler, R. N. Fruit From the Sands: The Silk Road Origins of the Foods We Eat (University of California Press, 2019).
Miller, N. F., Spengler, R. N. & Frachetti, M. Millet cultivation across Eurasia: Origins, spread, and the influence of seasonal climate. The Holocene 26, 1566–1575 (2016).
Zohary, D., Hopf, M. & Weiss, E. Domestication of Plants in the Old World: The Origin and Spread of Cultivated Plants in West Asia, Europe and the Nile Valley (Oxford University Press, 2012).
Amadou, I., Gounga, M. E. & Le, G.-W. Millets: Nutritional composition, some health benefits and processing: A review. Emir. J. Food Agric. 25, 501–508 (2013).
Lyon, D. J. et al. Producing and Marketing Proso Millet in the Great Plains. Univ. Neb. Ext. Circ. #EC137 (2008).
Rachie, K. O. The Millets: Importance, Utilization and Outlook (International Crops Research Institute for the Semi-Arid Tropics (ICRISAT), 1975).
Fuller, D. Q., Boivin, N., Hoogervorst, T. & Allaby, R. Across the Indian Ocean: The prehistoric movement of plants and animals. Antiquity 85, 544–558 (2011).
Jones, M. et al. Food globalisation in prehistory: The agrarian foundations of an interconnected continent. J. Br. Acad. 4, 73–87 (2016).
Jones, M. et al. Food globalization in prehistory. World Archaeol. 43, 665–675 (2011).
Liu, X. et al. From ecological opportunism to multi-cropping: Mapping food globalisation in prehistory. Quat. Sci. Rev. 206, 21–28 (2019).
Sherratt, A. The Trans-Eurasian exchange: The prehistory of Chinese relations with the West. In Contact and Exchange in the Ancient World (ed. Mair, V. H.) 30–53 (University of Hawaii Press, 2006).
Wirth, E. Agrargeographie des Irak (Instituts fur Geographie und Wirtschaftsgeographie der Universitat Hamburg, 1962).
FAO/GIEWS. FAO GIEWS Country Brief on Iraq. http://www.fao.org/giews/countrybrief/country.jsp?lang=en&code=IRQ (2020).
Bor, N. L. Gramineae. In Flora of Iraq, Gramineae Vol. 9 (eds Townsend, C. C. et al.) (Ministry of Agriculture & Agrarian Reform, 1968).
Rosenzweig, M. S. ‘Ordering the chaotic periphery’: The environmental impact of the neo-assyrian empire on its provinces. In The Provincial Archaeology of the Assyrian Empire (eds MacGinnis, J. et al.) 49–58 (Oxbow Press, 2016).
Fuller, D. Q. & Boivin, N. Crops, cattle and commensals across the Indian Ocean: Current and Potential Archaeobiological Evidence. Études Océan Indien 42–43, 13–46 (2009).
Barjamovic, G. Interlocking commercial networks and the infrastructure of trade in western asia during the bronze age. In Trade and Civilisation: Economic Networks and Cultural Ties from Prehistory to the Early Modern Era (eds Kristiansen, K. et al.) 113–142 (Cambridge University Press, 2018).
Frachetti, M. D., Smith, C. E., Traub, C. M. & Williams, T. Nomadic ecology shaped the highland geography of Asia’s Silk Roads. Nature 543, 193–198 (2017).
Spengler, R. et al. Early agriculture and crop transmission among Bronze Age mobile pastoralists of Central Eurasia. Proc. R. Soc. B Biol. Sci. 281, 20133382 (2014).
Fick, S. E. & Hijmans, R. J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315 (2017).
Williams, T. The Silk Roads: An ICOMOS Thematic Study (ICOMOS, 2014).
Charles, M. Introductory remarks on the cereals. In Bulletin on Sumerian Agriculture Vol. 1 (eds Postgate, J. N. & Powell, M. A.) 17–31 (University of Cambridge, 1984).
Powell, M. A. Sumerian cereal crops. In Bulletin on Sumerian Agriculture Vol. 1 (eds Postgate, J. N. & Powell, M. A.) 48–72 (University of Cambridge, 1984).
The Assyrian Dictionary of the Oriental Institute of the University of Chicago: D. (ed. Oppenheim A.L. et al.) vol. 3 (Oriental Institute of the University of Chicago, 1959).
Widell, M. et al. Staple production, cultivation and sedentary life: Model Input data. In Models of Mesopotamian Landscapes: How Small-Scale Processes Contributed to the Growth of Early Civilizations (eds Wilkinson, T. J. et al.) 81–101 (Archaeopress, 2013).
Bedigian, D. Is še-giš-ì Sesame or Flax? In Bulletin on Sumerian Agriculture Vol. 2 (eds Postgate, J. N. & Powell, M. A.) 158–178 (University of Cambridge, 1985).
Waetzoldt, H. Ölpflanzen und Pflanzenöle im 3. Jahrtausend. In Bulletin on Sumerian Agriculture Vol. 2 (eds Postgate, J. N. & Powell, M. A.) 77–95 (University of Cambridge, 1985).
Maekawa, K. Cereal Cultivation in the Ur III period. In Bulletin on Sumerian Agriculture Vol. 1 (eds Postgate, J. N. & Powell, M. A.) 73–96 (University of Cambridge, 1984).
Nesbitt, M. & Summers, G. D. Some recent Discoveries of Millet (Panicum Miliaceum L. and Setaria italica (L.) P. Beauv.) at Excavations in Turkey and Iran. Anatol. Stud. 38, 85–97 (1988).
Charles, M. Botanical remains. In Abu Salabikh Excavations: The 6G Ash-Tip and Its Contents: Cultic and Administrative Discard from the Temple? Vol. 4 (ed. Green, A.) 203–207 (British School of Archaeology in Iraq, 1993).
Postgate, J. N. Early Mesopotamia: Society and Economy at the Dawn of History (Routledge, 1992).
Bulletin on Sumerian Agriculture. (eds. Postgate, J. N. & Powell, M. A.) vol. 1 (University of Cambridge, 1984).
Lawrence, D., Philip, G., Hunt, H., Snape-Kennedy, L. & Wilkinson, T. J. Long term population, city size and climate trends in the fertile crescent: A first approximation. PLoS ONE 11, e0152563 (2016).
Models of Mesopotamian landscapes: How small-scale processes contributed to the growth of early civilizations. (eds. Wilkinson, T.J., Gibson, M., & Widell, M.) (Archaeopress, 2013).
Charles, M. & Dobney, K. Mesopotamian Environmental Archaeology Database: Phase I Iraq . (Archaeology Data Service [distributor], 2009).
Helbaek, H. The Plant Remains from Nimrud. In Nimrud and its Remains Vol. II (ed. Mallowan, M. E. L.) 613–620 (Collins, 1966).
Boserup, E. The Conditions of Agricultural Growth: The Economics of Agrarian Change Under Population Pressure (Aldine Publishing Co., 1965).
Brite, E. B., Kidd, F. J., Betts, A. & Negus Cleary, M. Millet cultivation in Central Asia: A response to Miller et al.. The Holocene 27, 1415–1422 (2017).
Helbaek, H. The Paleoethnobotany of the Near East and Europe. In Prehistoric Investigations in Iraqi Kurdistan (eds Braidwood, R. J. & Howe, B.) 99–118 (University of Chicago Press, 1960).
Jacobsen, T. Salinity and Irrigation Agriculture in Antiquity: Diyala Basin Archaeological Projects: Report on Essential Results, 1957–58 (Undena Publ, 1982).
Motuzaite-Matuzeviciute, G., Staff, R. A., Hunt, H. V., Liu, X. & Jones, M. K. The early chronology of broomcorn millet (Panicum miliaceum) in Europe. Antiquity 87, 1073–1085 (2013).
Field, H. Ancient Wheat and Barley from Kish, Mesopotamia. Am. Anthropol. 34, 303–309 (1932).
Wengrow, D. et al. Gurga Chiya and Tepe Marani: New Excavations in the Shahrizor Plain, Iraqi Kurdistan. Iraq 78, 253–284 (2016).
van Zeist, W. Comments on Plant Cultivation at Two Sites on the Khabur, North-Eastern Syria. In Umwelt und Subsistenze der assyrischen Stadt Dur-Katlimmu am unteren Habur Vol. 8 (ed. Kühne, H.) 133–147 (Harrassowitz, 2008).
van Zeist, W. Comments on plant cultivation at two sites on the Khabur, North-eastern Syria. in Reports on archaeobotanical studies in the Old World. (ed. van Zeist, W.) 33–60 (2003).
Riehl, S. Erste ergebnisse der archäobotanischen untersuchungen in der zentralen oberstadt von Tall Mozan/Urkeš im rahmen der DOG-IIMAS-Kooperation. Mitteilungen Dtsch. Orient-Ges. Zu Berl. 132, 229–238 (2000).
Riehl, S. Plant production in a changing environment: The archaeobotanical remains from Tell Mozan. In Development of the Environment, Subsistence and Settlement of the City of Urkeš and Its Region (eds Deckers, K. et al.) 13–158 (Harrassowitz, 2010).
Glatz, C. et al. Babylonian Encounters in the Upper Diyala Valley: Contextualizing the Results of Regional Survey and the 2016–2017 Excavations at Khani Masi. Am. J. Archaeol. 123, 439–471 (2019).
Glatz, C. & Casana, J. Of highland-lowland borderlands: Local societies and foreign power in the zagros-mesopotamian interface. J. Anthropol. Archaeol. 44, 127–147 (2016).
Perruchini, E., Glatz, C., Hald, M. M., Casana, J. & Toney, J. L. Revealing invisible brews: A new approach to the chemical identification of ancient beer. J. Archaeol. Sci. 100, 176–190 (2018).
Laugier, E. J., Casana, J., Glatz, C., Sameen, S. M. & Cabanes, D. Reconstructing agro-pastoral practice in the Mesopotamian-Zagros borderlands: Insights from phytolith and FTIR analysis of a dung-rich deposit. J. Archaeol. Sci. Rep. 38, 103106 (2021).
Zohary, M. Geobotanical foundations of the Middle East (Fischer, 1973).
Sinha, A. et al. Role of climate in the rise and fall of the Neo-Assyrian Empire. Sci. Adv. 5, eaax6656 (2019).
Schneider, U., Becker, A., Finger, P., Rustemeier, E. & Ziese, M. GPCC Full Data Monthly Product Version 2020 at 0.25°: Monthly Land-Surface Precipitation from Rain-Gauges built on GTS-based and Historical Data. (2020).
Casana, J. & Glatz, C. The land behind the land behind baghdad: Archaeological landscapes of the upper Diyala (Sirwan) River Valley. Iraq 79, 1–23 (2017).
Wilkinson, T. J. Settlement and land use in the zone of uncertainty in upper mesopotamia. In Rainfall and Agriculture in Northern Mesopotamia (ed. Jas, R. M.) 3–35 (Nederlands Historisch-Archaeologisch Instituut te Istanbul, 2000).
Saleh, D. K. Stream gage descriptions and streamflow statistics for sites in the Tigris River and Euphrates River basins, Iraq. (US Department of the Interior, US Geological Survey Reston, VA, USA, 2010).
Bar-Oz, G. et al. Ancient trash mounds unravel urban collapse a century before the end of Byzantine hegemony in the southern Levant. Proc. Natl. Acad. Sci. 116, 8239–8248 (2019).
Shillito, L.-M. & Matthews, W. Geoarchaeological investigations of midden-formation processes in the early to late ceramic neolithic levels at Çatalhöyük, Turkey ca. 8550–8370 cal BP. Geoarchaeology 28, 25–49 (2013).
McCorriston, J. & Weisberg, S. Spatial and temporal variation in mesopotamian agricultural practices in the Khabur Basin, Syrian Jazira. J. Archaeol. Sci. 29, 485–498 (2002).
Stone, E. C. Nippur neighborhoods (The Oriental Institute of the University of Chicago, 1987).
Bronk Ramsey, C. OxCal v4.4.4. (2021).
Madella, M., Alexandre, A. & Ball, T. International code for phytolith nomenclature 1.0. Ann. Bot. 96, 253–260 (2005).
Neumann, K. et al. International code for phytolith nomenclature (ICPN) 2.0. Ann. Bot. 20, 189–199 (2019).
Ge, Y., Lu, H., Zhang, J., Wang, C. & Gao, X. Phytoliths in inflorescence bracts: Preliminary results of an investigation on common panicoideae plants in China. Front. Plant Sci. 10, 1736 (2020).
Parry, D. W. & Hodson, M. J. Silica distribution in the caryopsis and inflorescence bracts of foxtail millet [Setaria italica (L.) Beauv.] and its Possible Significance in Carcinogenesis. Ann. Bot. 49, 531–540 (1982).
Cabanes, D. et al. Human impact around settlement sites: A phytolith and mineralogical study for assessing site boundaries, phytolith preservation, and implications for spatial reconstructions using plant remains. J. Archaeol. Sci. 39, 2697–2705 (2012).
Cabanes, D. & Shahack-Gross, R. Understanding fossil phytolith preservation: The role of partial dissolution in paleoecology and archaeology. PLoS ONE 10, e0125532 (2015).
Li, Z., de Tombeur, F., Linden, C. V., Cornelis, J.-T. & Delvaux, B. Soil microaggregates store phytoliths in a sandy loam. Geoderma 360, 114037 (2020).
Goldberg, P. & Macphail, R. I. Practical and Theoretical Geoarchaeology (Blackwell Publishing Ltd., 2006).
Ball, T. B. et al. Phytoliths as a tool for investigations of agricultural origins and dispersals around the world. J. Archaeol. Sci. 68, 32–45 (2016).
Kealhofer, L., Huang, F., DeVincenzi, M. & Kim, M. M. Phytoliths in Chinese foxtail millet (Setaria italica). Rev. Palaeobot. Palynol. 223, 116–127 (2015).
Weisskopf, A. R. & Lee, G.-A. Phytolith identification criteria for foxtail and broomcorn millets: A new approach to calculating crop ratios. Archaeol. Anthropol. Sci. 8, 29–42 (2016).
Lu, H. et al. Phytoliths analysis for the discrimination of foxtail millet (Setaria italica) and common millet (Panicum miliaceum). PLoS ONE 4, e4448 (2009).
Zhang, J. et al. Phytolith analysis for differentiating between broomcorn millet (Panicum miliaceum) and its weed/feral type (Panicum ruderale). Sci. Rep. 8, 1–9 (2018).
Nesbitt, M. Identification Guide for Near Eastern Grass Seeds (Institute of Archaeology, UCL, 2006).
Rudov, A., Mashkour, M., Djamali, M. & Akhani, H. A review of C4 plants in southwest asia: An ecological, geographical and taxonomical analysis of a region with high diversity of C4 eudicots. Front. Plant Sci. 11, 1374 (2020).
Weber, S. A. & Fuller, D. Q. Millets and their role in early agriculture. Pragdhara 18, 69–90 (2008).
Zhang, J., Lu, H., Wu, N., Yang, X. & Diao, X. Phytolith analysis for differentiating between foxtail millet (Setaria italica) and green foxtail (Setaria viridis). PLoS ONE 6, e19726 (2011).
Out, W. A. & Madella, M. Morphometric distinction between bilobate phytoliths from Panicum miliaceum and Setaria italica leaves. Archaeol. Anthropol. Sci. 8, 505–521 (2016).
Bhat, M. A., Shakoor, S. A., Badgal, P. & Soodan, A. S. Taxonomic Demarcation of Setaria pumila (Poir.) Roem. & Schult., S. verticillata (L.) P. Beauv., and S. viridis (L.) P. Beauv. (Cenchrinae, Paniceae, Panicoideae, Poaceae) From Phytolith Signatures. Front. Plant Sci. 9, 864 (2018).
Ge, Y. et al. Phytolith analysis for the identification of barnyard millet (Echinochloa sp.) and its implications. Archaeol. Anthropol. Sci. 10, 61–73 (2018).
Madella, M., Lancelotti, C. & García-Granero, J. J. Millet microremains—an alternative approach to understand cultivation and use of critical crops in Prehistory. Archaeol. Anthropol. Sci. 8, 17–28 (2016).
Ball, T. B., Vrydaghs, L., Van Den Hauwe, I., Manwaring, J. & De Langhe, E. Differentiating banana phytoliths: Wild and edible Musa acuminata and Musa balbisiana. J. Archaeol. Sci. 33, 1228–1236 (2006).
Ball, T. B. et al. Morphometric analysis of phytoliths: Recommendations towards standardization from the International Committee for Phytolith Morphometrics. J. Archaeol. Sci. 68, 106–111 (2016).
Hunt, H. V. et al. Millets across Eurasia: Chronology and context of early records of the genera Panicum and Setaria from archaeological sites in the Old World. Veg. Hist. Archaeobotany 17, 5 (2008).
Weisskopf, A. R. Millets, Rice and Farmers: Phytoliths as Indicators of Agricultural, Social and Ecological Change in Neolithic and Bronze Age Central China (British Archaeological Reports, 2014).
Ghazanfar, S. A. & McDaniel, T. Floras of the Middle East: A Quantitative Analysis and Biogeography of the Flora of Iraq. Edinb. J. Bot. 73, 1–24 (2016).
Reddy, S. N. If the threshing floor could talk: Integration of agriculture and pastoralism during the late harappan in Gujarat, India. J. Anthropol. Archaeol. 16, 162–187 (1997).
Liu, X. & Jones, M. K. Food globalisation in prehistory: Top down or bottom up?. Antiquity 88, 956–963 (2014).
Adams, RMc. C. Heartland of Cities: Surveys of Ancient Settlement and Land Use on the Central Floodplain of the Euphrates (University of Chicago Press, 1981).
Oates, J. Choga Mami, 1967–68: A Preliminary Report. Iraq 31, 115–152 (1969).
Rost, S. Navigating the ancient Tigris: Insights into water management in an early state. J. Anthropol. Archaeol. 54, 31–47 (2019).
Dunseth, Z. C. et al. Archaeobotanical proxies and archaeological interpretation: A comparative study of phytoliths, pollen and seeds in dung pellets and refuse deposits at Early Islamic Shivta, Negev, Israel. Quat. Sci. Rev. 211, 166–185 (2019).
Amarna Diplomacy: The Beginnings of International Relations. (Johns Hopkins Univeristy Press, 2000).
Kenoyer, J. M. Indus and Mesopotamian Trade Networks: New Insights from Shell and Carnelian Artifacts. In Intercultural Relations Between South and Southwest Asia. Studies In Commemoration Of E.C.L.During Caspers (1934–1996) (eds Olijdam, E. & Spoor, R. H.) 19–28 (Archaeopress, 2008).
Scott, A. et al. Exotic foods reveal contact between South Asia and the Near East during the second millennium BCE. Proc. Natl. Acad. Sci. 118, 1–10 (2020).
Schiffer, M. B. Formation Processes of the Archaeological Record (University of New Mexico Press, 1987).
Bates, J., Singh, R. N. & Petrie, C. A. Exploring Indus crop processing: Combining phytolith and macrobotanical analyses to consider the organisation of agriculture in northwest India c. 3200–1500 bc. Veg. Hist. Archaeobotany 26, 25–41 (2017).
Harvey, E. L. & Fuller, D. Q. Investigating crop processing using phytolith analysis: The example of rice and millets. J. Archaeol. Sci. 32, 739–752 (2005).
Hillman, G. Interpretation of archaeological plant remains: The application of ethnographic models from Turkey. in Plants and Ancient Man: Studies in Palaeoethnobotany: Proceedings of the Sixth Symposium of the International Work Group for Palaeoethnobotany, Groningen, 30 May-3 June 1983 (eds. van Zeist, W. & Casparie, W. A.) 1–41 (Balkema, 1984).
Hillman, G. Reconstructing crop husbandry practices from charred remains of crops. In Farming Practice in British prehistory (ed. Mercer, R.) 123–162 (Edinburgh University Press, 1981).
Helbaek, H. Samarran irrigation agriculture at Choga Mami in Iraq. Iraq 34, 35–48 (1972).
Miller, N. F. Economy and Environment of Malyan, a Third Millennium BC Urban Center in Southern Iran (The University of Michigan, 1982).
Bedigian, D. Sesame: The Genus Sesamum (CRC Press, 2010).
Van Zeist, W. Some notes on second millennium BC plant cultivation in the Syrian Jazira. In Cinquante-deux réflexions sur le Proche-Orient ancien offertes en hommage a Leon de Meijer (eds Gasche, H. & Tanret, M.) 541–553 (Peeters, 1994).
Linares, V. et al. First evidence for vanillin in the old world: Its use as mortuary offering in Middle Bronze Canaan. J. Archaeol. Sci. Rep. 25, 77–84 (2019).
Chowdhury, M. P., Campbell, S. & Buckley, M. Proteomic analysis of archaeological ceramics from Tell Khaiber, southern Iraq. J. Archaeol. Sci. 132, 105414 (2021).
Miller, N. & Smart, T. Intentional burning of dung as fuel: A mechanism for the incorporation of charred seeds into the archaeological record. J. Ethnobiol. 4, 15–28 (1984).
Putman, R. J. Facts from faeces. Mammal Rev. 14, 79–97 (1984).
Shahack-Gross, R. Herbivorous livestock dung: Formation, taphonomy, methods for identification, and archaeological significance. J. Archaeol. Sci. 38, 205–218 (2011).
Valamoti, S. M. & Charles, M. Distinguishing food from fodder through the study of charred plant remains: An experimental approach to dung-derived chaff. Veg. Hist. Archaeobotany 14, 528–533 (2005).
Wallace, M. & Charles, M. What goes in does not always come out: The impact of the ruminant digestive system of sheep on plant material, and its importance for the interpretation of dung-derived archaeobotanical assemblages. Environ. Archaeol. 18, 18–30 (2013).
Hammer, E. L. & Arbuckle, B. S. 10,000 Years of pastoralism in anatolia: A review of evidence for variability in pastoral lifeways. Nomadic Peoples 21, 214–267 (2017).
Meadow, R. H. Inconclusive remarks on pastoralism, nomadism, and other animal-related matters. In Pastoralism in the Levant: Archaeological Materials in Anthropological Perspectives (eds Bar-Yosef, O. & Khazanov, A. M.) 261–269 (Prehistory Press, 1992).
Frachetti, M. D. Multiregional emergence of mobile pastoralism and nonuniform institutional complexity across Eurasia. Curr. Anthropol. 53, 2–38 (2012).
García-Granero, J. J., Lancelotti, C., Madella, M. & Ajithprasad, P. Millets and herders: The origins of plant cultivation in semiarid North Gujarat (India). Curr. Anthropol. 57, 149–173 (2016).
Hermes, T. R. et al. Early integration of pastoralism and millet cultivation in Bronze Age Eurasia. Proc. R. Soc. B Biol. Sci. 286, 20191273 (2019).
Lightfoot, E., Liu, X. & Jones, M. K. Why move starchy cereals? A review of the isotopic evidence for prehistoric millet consumption across Eurasia. World Archaeol. 45, 574–623 (2013).
Miller, A. R. V. & Makarewicz, C. A. Intensification in pastoralist cereal use coincides with the expansion of trans-regional networks in the Eurasian Steppe. Sci. Rep. 9, 1–12 (2019).
Spengler, R. N., Frachetti, M. D. & Fritz, G. J. Ecotopes and herd foraging practices in the steppe/mountain ecotone of Central Asia during the Bronze and Iron Ages. J. Ethnobiol. 33, 125–147 (2013).
Arbuckle, B. S. & Hammer, E. L. The rise of pastoralism in the ancient near east. J. Archaeol. Res. 27, 391–449 (2019).
Paulette, T. Grain, Storage, and State Making in Mesopotamia (3200–2000 BC). In Storage in Ancient Complex Societies: Administration, Organization, and Control (eds Manzanilla, L. & Rothman, M. S.) 85–109 (Routledge, 2016).
Scott, J. C. Against the Grain: A Deep History of the Earliest States (Yale University Press, 2017).
Prehistoric Archeology along the Zagros Flanks. (eds. Braidwood, L.S., et al.) (Oriental Institute of the University of Chicago, 1983).
Liu, X., Hunt, H. V. & Jones, M. K. River valleys and foothills: Changing archaeological perceptions of North China’s earliest farms. Antiquity 83, 82–95 (2009).
Marston, J. M. Modeling resilience and sustainability in ancient agricultural systems. J. Ethnobiol. 35, 585–605 (2015).
Sołtysiak, A. & Schutkowski, H. Stable isotopic evidence for land use patterns in the Middle Euphrates Valley, Syria. Am. J. Phys. Anthropol. 166, 861–874 (2018).
Marston, J. M. Archaeological markers of agricultural risk management. J. Anthropol. Archaeol. 30, 190–205 (2011).
Katz, O. et al. Rapid phytolith extraction for analysis of phytolith concentrations and assemblages during an excavation: An application at Tell es-Safi/Gath, Israel. J. Archaeol. Sci. 37, 1557–1563 (2010).
Piperno, D. R. Phytoliths: A Comprehensive Guide for Archaeologists and Paleoecologists (AltaMira Press, 2006).
Piperno, D. R. Phytolith Analysis: An Archaeological and Geological Perspective (Academic Press, 1988).
Phytolith Systematics: Emerging Issues. (eds. Rapp, G. & Mulholland, S.C.) vol. 1 (Springer, 1992).
Twiss, P. C., Suess, E. & Smith, R. M. Morphological classification of grass phytoliths. Soil Sci. Soc. Am. J. 33, 109–115 (1969).
Acknowledgements
This study was supported by the Ecology, Evolution, Environment, and Society (EEES) Program and the Department of Anthropology’s Claire Garber Goodman Fund at Dartmouth College. Fieldwork at Khani Masi, including radiocarbon dates, was funded by the National Science Foundation (1724488) and was conducted in collaboration with the Garmian Directorate of Antiquities with the permission of the General Directorate of Antiquities of the Kurdistan Region of Iraq. We thank Kaify Ali Mustafa, Director General of Antiquities for the Kurdistan Region, and Salah Mohammed Sameen, Deputy Director of the Garmian Museum, for their ongoing support; SRP co-director, Claudia Glatz, the SRP field team, and Museum Garmian Museum representatives who conducted the excavations; Nathaniel Dominy for the use of his laboratory space; and Carla Lancelotti and Alexia Smith for helpful comments on the manuscript.
Author information
Authors and Affiliations
Contributions
E.J.L., J.C., and D.C. designed research; E.J.L. performed research; E.J.L., J.C., and D.C. analyzed data; E.J.L. wrote the paper; and J.C. and D.C. contributed to writing of the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Laugier, E.J., Casana, J. & Cabanes, D. Phytolith evidence for the pastoral origins of multi-cropping in Mesopotamia (ancient Iraq). Sci Rep 12, 60 (2022). https://doi.org/10.1038/s41598-021-03552-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-03552-w
This article is cited by
-
Pre-Aksumite plant husbandry in the Horn of Africa
Vegetation History and Archaeobotany (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
