Preparation and Optimisation of Cross-Linked Enzyme Aggregates Using Native Isolate White Rot Fungi Trametes versicolor and Fomes fomentarius for the Decolourisation of Synthetic Dyes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Laccase Production
2.2. Optimisation of CLEAs Preparation Conditions
2.3. Laccase Activity and Protein Assay
2.4. pH Optimum and Stability of Free and Immobilised Laccase
2.5. Optimal Temperature and Thermal Stability of Free and Immobilised Laccase
2.6. Storage and Operational Stability of Free and Immobilised Laccase
2.7. Stability of Free and Immobilised Laccase against Denaturation
2.8. Kinetic Parameters of Free and Immobilised Laccase
2.9. Decolourisation Experiment
3. Results and Discussion
3.1. Optimisation of CLEA Preparation
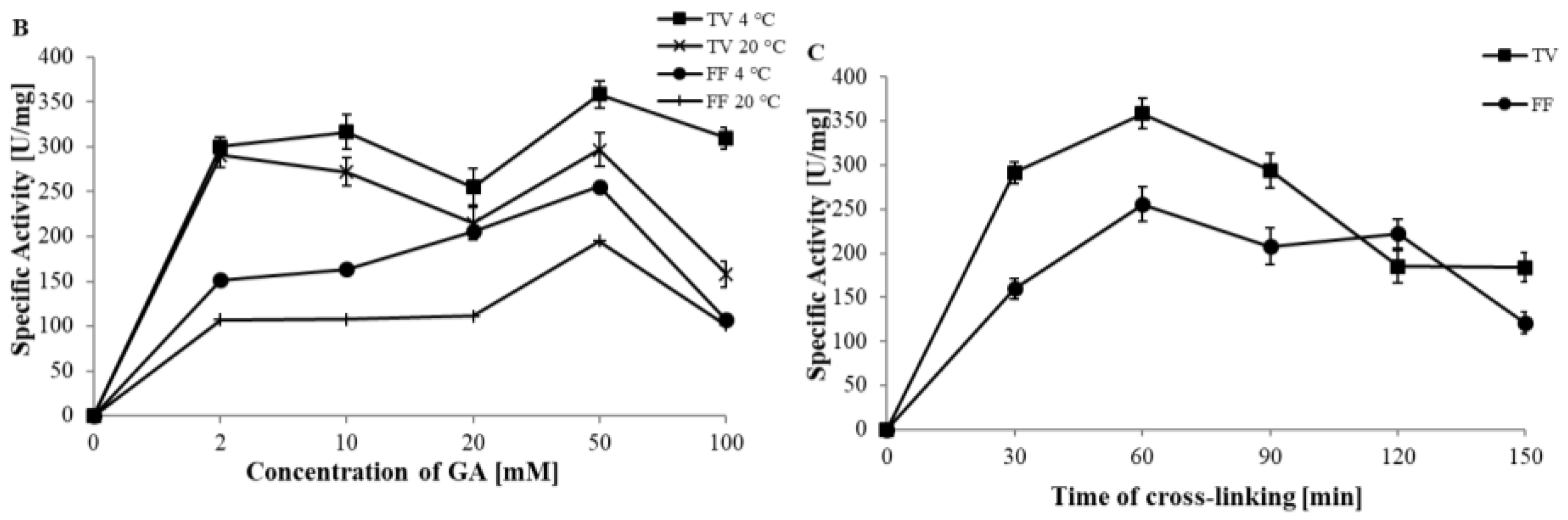
3.1.1. The Effect of Cross-Linking Time
3.1.2. Effects of Cross-Linking Temperature, Time, and Different Concentrations of Cross-Linking Agent
3.2. Characterisation of CLEA Laccase
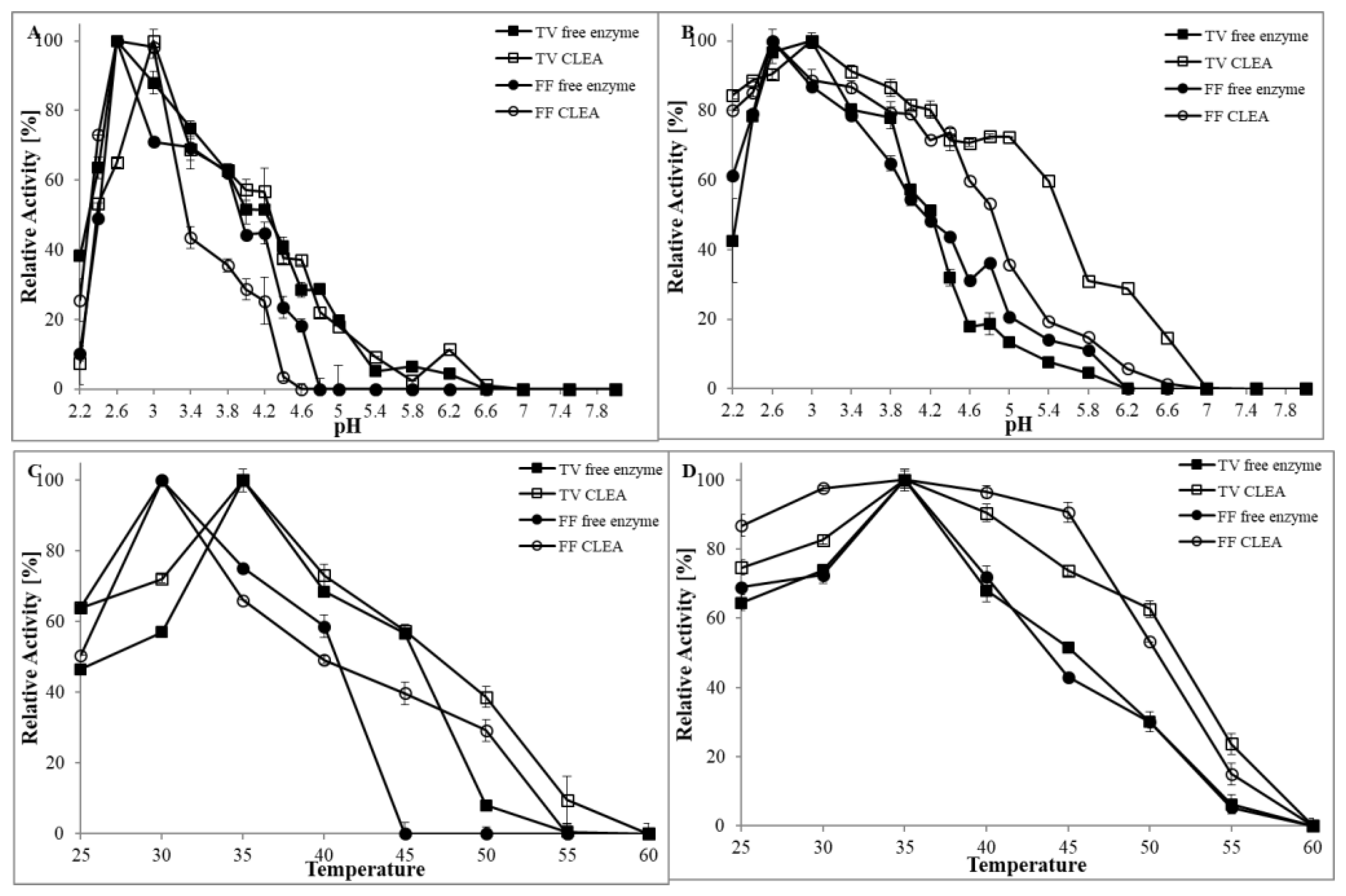
3.2.1. The Influence of pH
3.2.2. The Influence of Temperature
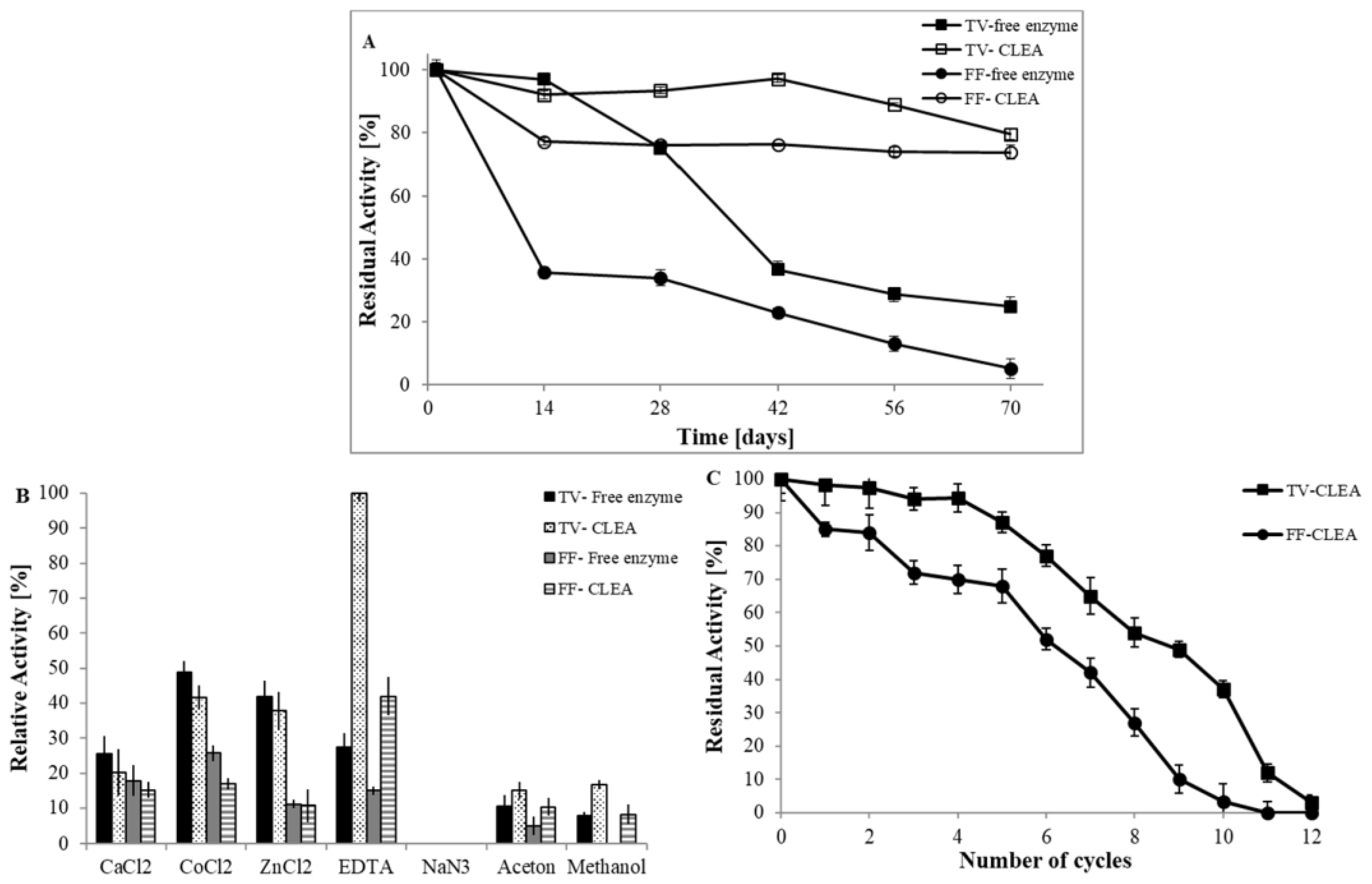
3.2.3. Storage Stability
3.2.4. Stability against Denaturing Conditions
3.2.5. Kinetic Parameters of Free and Immobilised Laccase
3.2.6. Operational Stability of Immobilised Laccase
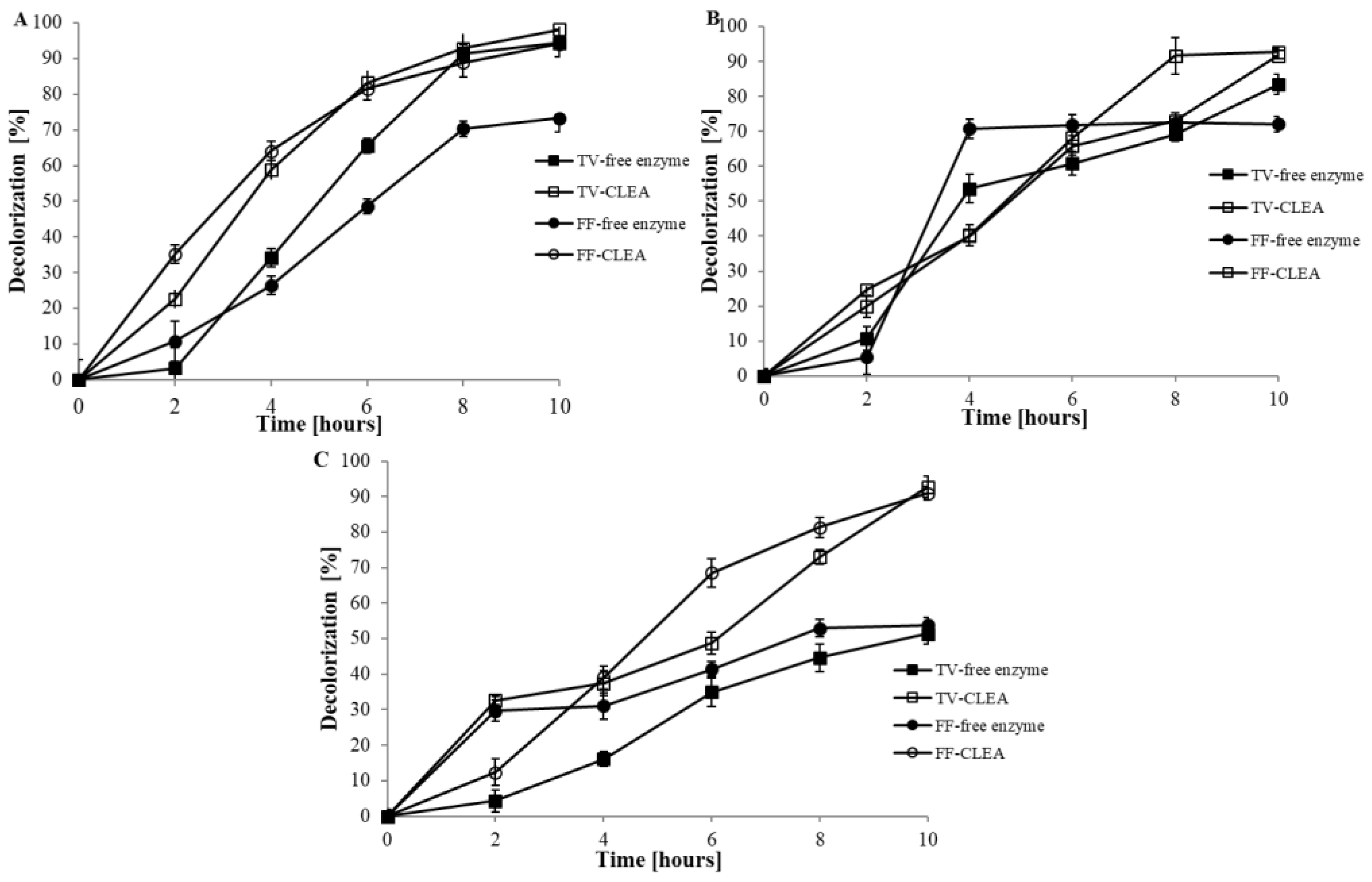
3.2.7. Decolourisation Experiment
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Worrall, J.J.; Anagnost, S.E.; Zabel, R.A. Comparison of wood decay among diverse lignicolous fungi. Mycologia 1997, 89, 199–219. [Google Scholar] [CrossRef]
- Giardina, P.; Faraco, V.; Pezzella, C.; Piscitelli, A.; Vanhulle, S.; Sannia, G. Laccases: A never-ending story. Cell. Mol. Life Sci. 2010, 67, 369–385. [Google Scholar] [CrossRef] [PubMed]
- Nyanhongo, G.S.; Gomes, J.; Gubitz, G.M.; Zvauya, R.; Read, J.; Steiner, W. Decolorization of textile dyes by laccases from a newly isolated strain of Trametes modesta. Water Res. 2002, 36, 1449–1456. [Google Scholar] [CrossRef]
- Baldrian, P. Fungal laccases-occurrence and properties. FEMS Microbiol. Rev. 2006, 30, 215–242. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, B.; Martinez-Morales, F.; Trejo-Hernandez, M.R. Fungal laccases: Induction and production. Rev. Mex. Ing. Quim. 2013, 12, 473–488. [Google Scholar]
- Viswanath, B.; Rajesh, B.; Janardhan, A.; Kumar, A.P.; Narasimha, G. Fungal laccases and their applications in bioremediation. Enzyme Res. 2014, 2014, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Bosso, L.; Cristinzio, G. A comprehensive overview of bacteria and fungi used for pentachlorophenol biodegradation. Rev. Environ. Sci. Bio/Technol. 2014, 13, 387–427. [Google Scholar] [CrossRef]
- McMullan, G.; Meehan, C.; Conneely, A.; Kirby, N.; Robinson, T.; Nigam, P.; Banat, I.M.; Marchant, R.; Smyth, W.E. Microbial decolourisation and degradation of textile dyes. Appl. Microbiol. Biotechnol. 2001, 56, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Maciel, M.J.M.; Silva, A.C.E.; Ribeiro, H.C.T. Industrial and biotechnological applications of ligninolytic enzymes of the basidiomycota: A review. Electron. J. Biotechnol. 2010, 13, 1–13. [Google Scholar]
- Matijosyte, I.; Arends, I.; de Vries, S.; Sheldon, R.A. Preparation and use of cross-linked enzyme aggregates (cleas) of laccases. J. Mol. Catal. B Enzym. 2010, 62, 142–148. [Google Scholar] [CrossRef]
- Talekar, S.; Joshi, A.; Joshi, G.; Kamat, P.; Haripurkar, R.; Kambale, S. Parameters in preparation and characterization of cross linked enzyme aggregates (cleas). RSC Adv. 2013, 3, 12485–12511. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Pletschke, B.I. Magnetic cross-linked enzyme aggregates (cleas): A novel concept towards carrier free immobilization of lignocellulolytic enzymes. Enzyme Microb. Technol. 2014, 61–62, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.V.; Cabana, H. Towards high potential magnetic biocatalysts for on-demand elimination of pharmaceuticals. Bioresour. Technol. 2016, 200, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Talekar, S.; Ghodake, V.; Ghotage, T.; Rathod, P.; Deshmukh, P.; Nadar, S.; Mulla, M.; Ladole, M. Novel magnetic cross-linked enzyme aggregates (magnetic cleas) of alpha amylase. Bioresour. Technol. 2012, 123, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, N.R.; Marzuki, N.H.C.; Buang, N.A.; Huyop, F.; Wahab, R.A. An overview of technologies for immobilization of enzymes and surface analysis techniques for immobilized enzymes. Biotechnol. Biotechnol. Equip. 2015, 29, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A. Enzyme immobilization: The quest for optimum performance. Adv. Synth. Catal. 2007, 349, 1289–1307. [Google Scholar] [CrossRef]
- Sheldon, R.A. Cross-linked enzyme aggregates as industrial biocatalysts. Org. Process Res. Dev. 2011, 15, 213–223. [Google Scholar] [CrossRef]
- Sheldon, R.A.; van Pelt, S. Enzyme immobilisation in biocatalysis: Why, what and how. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.L.; Xian, H.; Shi, S.J.; Zhang, C.S.; Manik, S.M.N.; Mao, J.J.; Zhang, G.; Liao, W.H.; Wang, Q.; Liu, H.B. Biodegradation of lignin and nicotine with white rot fungi for the delignification and detoxification of tobacco stalk. BMC Biotechnol. 2016, 16, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.V.; Sivanesan, S.; Cabana, H. Magnetic cross-linked laccase aggregates—Bioremediation tool for decolorization of distinct classes of recalcitrant dyes. Sci. Total Environ. 2014, 487, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zou, M.; Hong, F.F. Evaluation of fungal laccase immobilized on natural nanostructured bacterial cellulose. Front. Microbiol. 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Alqadami, A.A.; Naushad, M.; Abdalla, M.A.; Khan, M.R.; Alothman, Z.A. Adsorptive removal of toxic dye using Fe3O4-TSC nanocomposite: Equilibrium, kinetic, and thermodynamic studies. J. Chem. Eng. Data 2016, 61, 3806–3813. [Google Scholar] [CrossRef]
- Daneshvar, E.; Vazirzadeh, A.; Niazi, A.; Kousha, M.; Naushad, M.; Bhatnagar, A. Desorption of methylene blue dye from brown macroalga: Effects of operating parameters, isotherm study and kinetic modeling. J. Clean. Prod. 2017, 152, 443–453. [Google Scholar] [CrossRef]
- Pathania, D.; Katwal, R.; Sharma, G.; Naushad, M.; Khan, M.R.; Al-Muhtaseb, A.H. Novel guar gum/Al2O3 nanocomposite as an effective photocatalyst for the degradation of malachite green dye. Int. J. Biol. Macromol. 2016, 87, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Pathania, D.; Gupta, D.; Al-Muhtaseb, A.H.; Sharma, G.; Kumar, A.; Naushad, M.; Ahamad, T.; Alshehri, S.M. Photocatalytic degradation of highly toxic dyes using chitosan-g-poly (acrylamide)/ZNS in presence of solar irradiation. J. Photochem. Photobiol. A Chem. 2016, 329, 61–68. [Google Scholar] [CrossRef]
- Vrsanska, M.; Voberkova, S. The ability to decolorize different synthetic dyes due to laccase production by trametes versicolor and fomes fomentarius. In Proceedings of the International PhD Students Conference (Mendelnet 2016), Brno, Czech Republic, 9–10 November 2016; pp. 763–768. [Google Scholar]
- Solcany, V.; Vrsanska, M.; Voberkova, S. Optimization of the procedure for a ligninolytic enzymes isolation from the white rot fungi. In Proceedings of the International phD Students Conference (Mendelnet 2016), Brno, Czech Republic, 9–10 November 2016; pp. 1011–1015. [Google Scholar]
- Li, S.P.; Su, Y.T.; Liu, Y.D.; Sun, L.F.; Yu, M.X.; Wu, Y.K. Preparation and characterization of cross-linked enzyme aggregates (cleas) of recombinant thermostable alkylsulfatase (sdsap) from pseudomonas sp s9. Process Biochem. 2016, 51, 2084–2089. [Google Scholar] [CrossRef]
- Jiang, D.S.; Long, S.Y.; Huang, J.; Xiao, H.Y.; Zhou, J.Y. Immobilization of pycnoporus sanguineus laccase on magnetic chitosan microspheres. Biochem. Eng. J. 2005, 25, 15–23. [Google Scholar] [CrossRef]
- Kumar, V.V.; Kumar, M.P.P.; Thiruvenkadaravi, K.V.; Baskaralingam, P.; Kumar, P.S.; Sivanesan, S. Preparation and characterization of porous cross linked laccase aggregates for the decolorization of triphenyl methane and reactive dyes. Bioresour. Technol. 2012, 119, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Bourbonnais, R.; Paice, M.G. Oxidation of non-phenolic substrates: An expanded role for laccase in lignin biodegradation. FEBS Lett. 1990, 267, 99–102. [Google Scholar] [CrossRef]
- Wang, F.; Hu, J.H.; Guo, C.; Liu, C.Z. Enhanced laccase production by trametes versicolor using corn steep liquor as both nitrogen source and inducer. Bioresour. Technol. 2014, 166, 602–605. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Cristovao, R.O.; Silverio, S.C.; Tavares, A.P.M.; Brigida, A.I.S.; Loureiro, J.M.; Boaventura, R.A.R.; Macedo, E.A.; Coelho, M.A.Z. Green coconut fiber: A novel carrier for the immobilization of commercial laccase by covalent attachment for textile dyes decolourization. World J. Microbiol. Biotechnol. 2012, 28, 2827–2838. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Zeng, Z.T.; Zeng, G.M.; Tang, L.; Pang, Y.; Li, Z.; Liu, C.; Lei, X.X.; Wu, M.S.; Ren, P.Y.; et al. Immobilization of laccase on magnetic bimodal mesoporous carbon and the application in the removal of phenolic compounds. Bioresour. Technol. 2012, 115, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Kunamneni, A.; Ghazi, I.; Camarero, S.; Ballesteros, A.; Plou, F.J.; Alcalde, M. Decolorization of synthetic dyes by laccase immobilized on epoxy-activated carriers. Process Biochem. 2008, 43, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Davis, S.; Burns, R.G. Covalent immobilization of laccase on activated carbon for phenolic effluent treatment. Appl. Microbiol. Biotechnol. 1992, 37, 474–479. [Google Scholar] [CrossRef]
- Sinirlioglu, Z.A.; Sinirlioglu, D.; Akbas, F. Preparation and characterization of stable cross-linked enzyme aggregates of novel laccase enzyme from shewanella putrefaciens and using malachite green decolorization. Bioresour. Technol. 2013, 146, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Kastner, M.; Schulz, T. Ion-exchange fast protein liquid chromatography-optimization of the purification of cytochrome p 450 from marmoset monkeys. J. Chromatogr. 1987, 397, 153–163. [Google Scholar] [CrossRef]
- Lopez-Gallego, F.; Betancor, L.; Hidalgo, A.; Alonso, N.; Fernandez-Lafuente, R.; Guisan, J.M. Co-aggregation of enzymes and polyethyleneimine: A simple method to prepare stable and immobilized derivatives of glutaryl acylase. Biomacromolecules 2005, 6, 1839–1842. [Google Scholar] [CrossRef] [PubMed]
- Migneault, I.; Dartiguenave, C.; Bertrand, M.J.; Waldron, K.C. Glutaraldehyde: Behavior in aqueous solution, reaction with proteins, and application to enzyme crosslinking. Biotechniques 2004, 37, 790–796. [Google Scholar] [PubMed]
- Cabana, H.; Jones, J.P.; Agathos, S.N. Preparation and characterization of cross-linked laccase aggregates and their application to the elimination of endocrine disrupting chemicals. J. Biotechnol. 2007, 132, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Li, X.D.; Wu, J.; Jia, D.C.; Wan, Y.H.; Yang, N.; Qiao, M. Preparation of cross-linked glucoamylase aggregates immobilization by using dextrin and xanthan gum as protecting agents. Catalysts 2016, 6, 1. [Google Scholar] [CrossRef]
- Broun, G.B. Chemically aggregated enzymes. Methods Enzymol. 1976, 44, 263–280. [Google Scholar] [PubMed]
- Munoz, C.; Guillen, F.; Martinez, A.T.; Martinez, M.J. Laccase isoenzymes of pleurotus eryngii: Characterization, catalytic properties, and participation in activation of molecular oxygen and MN2+ oxidation. Appl. Environ. Microbiol. 1997, 63, 2166–2174. [Google Scholar] [PubMed]
- Wang, S.S.; Ning, Y.J.; Wang, S.N.; Zhang, J.; Zhang, G.Q.; Chen, Q.J. Purification, characterization, and cloning of an extracellular laccase with potent dye decolorizing ability from white rot fungus cerrena unicolor gsm-01. Int. J. Biol. Macromol. 2017, 95, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Xu, F. Effects of redox potential and hydroxide inhibition on the pH activity profile of fungal laccases. J. Biol. Chem. 1997, 272, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Neifar, M.; Jaouani, A.; Ellouze-Ghorbel, R.; Ellouze-Chaabouni, S. Purification, characterization and decolourization ability of fomes fomentarius laccase produced in solid medium. J. Mol. Catal. B Enzym. 2010, 64, 68–74. [Google Scholar] [CrossRef]
- Zhang, B.; Weng, Y.; Xu, H.; Mao, Z. Enzyme immobilization for biodiesel production. Appl. Microbiol. Biotechnol. 2012, 93, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Guo, C.; Yang, L.R.; Liu, C.Z. Magnetic mesoporous silica nanoparticles: Fabrication and their laccase immobilization performance. Bioresour. Technol. 2010, 101, 8931–8935. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Fernandez, M.; Sanroman, M.A.; Moldes, D. Recent developments and applications of immobilized laccase. Biotechnol. Adv. 2013, 31, 1808–1825. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Kaskel, S.; Shi, J.; Wage, T.; van Pee, K.H. Immobilization of trametes versicolor laccase on magnetically separable mesoporous silica spheres. Chem. Mater. 2007, 19, 6408–6413. [Google Scholar] [CrossRef]
- Ba, S.; Arsenault, A.; Hassani, T.; Jones, J.P.; Cabana, H. Laccase immobilization and insolubilization: From fundamentals to applications for the elimination of emerging contaminants in wastewater treatment. Crit. Rev. Biotechnol. 2013, 33, 404–418. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.F.; Jia, C.X.; Qi, W.; Yu, Q.X.; Peng, X.; Su, R.X.; He, Z.M. Porous-cleas of papain: Application to enzymatic hydrolysis of macromolecules. Bioresour. Technol. 2011, 102, 3541–3545. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, K.; Faramarzi, M.A.; Mahvi, A.H.; Gholami, M.; Esrafili, A.; Forootanfar, H.; Farzadkia, M. Elimination and detoxification of sulfathiazole and sulfamethoxazole assisted by laccase immobilized on porous silica beads. Int. Biodeterior. Biodegrad. 2015, 97, 107–114. [Google Scholar] [CrossRef]
- Bayramoglu, G.; Yilmaz, M.; Arica, M.Y. Preparation and characterization of epoxy-functionalized magnetic chitosan beads: Laccase immobilized for degradation of reactive dyes. Bioprocess Biosyst. Eng. 2010, 33, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.R.; Yin, L.F.; Niu, J.F. Laccase-carrying electrospun fibrous membranes for adsorption and degradation of PAHs in shoal soils. Environ. Sci. Technol. 2011, 45, 10611–10618. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Tang, R.Z.; Zhou, Q.J.; Li, F.T.; Zhang, B.R. Enhancement of catalytic activity of immobilized laccase for diclofenac biodegradation by carbon nanotubes. Chem. Eng. J. 2015, 262, 88–95. [Google Scholar] [CrossRef]
- Wilson, L.; Illanes, A.; Pessela, B.C.C.; Abian, O.; Fernandez-Lafuente, R.; Guisan, J.M. Encapsulation of crosslinked penicillin g acylase aggregates in lentikats: Evaluation of a novel biocatalyst in organic media. Biotechnol. Bioeng. 2004, 86, 558–562. [Google Scholar] [CrossRef] [PubMed]
- Sangeetha, K.; Abraham, T.E. Preparation and characterization of cross-linked enzyme aggregates (clea) of subtilisin for controlled release applications. Int. J. Biol. Macromol. 2008, 43, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Huang, J.; Ding, L.Y.; Li, M.T.; Chen, Z. Preparation of magnetic chitosan nanoparticles and immobilization of laccase. J. Wuhan Univ. Technol. Mater. Sci. Ed. 2009, 24, 42–47. [Google Scholar] [CrossRef]
- Ling, Z.R.; Wang, S.S.; Zhu, M.J.; Ning, Y.J.; Wang, S.N.; Li, B.; Yang, A.Z.; Zhang, G.Q.; Zhao, X.M. An extracellular laccase with potent dye decolorizing ability from white rot fungus trametes sp lac-01. Int. J. Biol. Macromol. 2015, 81, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Goel, G.; Sud, A.; Chauhan, R.S. A novel laccase from newly isolated cotylidia pannosa and its application in decolorization of synthetic dyes. Biocatal. Agric. Biotechnol. 2015, 4, 661–666. [Google Scholar] [CrossRef]
- Peralta-Zamora, P.; Pereira, C.M.; Tiburtius, E.R.L.; Moraes, S.G.; Rosa, M.A.; Minussi, R.C.; Duran, N. Decolorization of reactive dyes by immobilized laccase. Appl. Catal. B-Environ. 2003, 42, 131–144. [Google Scholar] [CrossRef]
- Rodríguez-Couto, S. Decolouration of industrial metal-complex dyes in successive batches by active cultures of trametes pubescens. Biotechnol. Rep. 2014, 4, 156–160. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration of glutaraldehyde (GA) (mM) | 2 | 10 | 20 | 50 | 100 |
| Time of cross-linking (min) | 30 | 60 | 90 | 120 | 150 |
| Temperature of cross-linking (°C) | 4 | 20 |
| Km Free Laccase (mM) | Km CLEA Laccase (mM) | Vmax Free Laccase (U/mg) | Vmax CLEA Laccase (U/mg) | |
|---|---|---|---|---|
| T. versicolor | 1.9 | 0.8 | 0.027 | 0.3 |
| F. fomentarius | 0.087 | 0.39 | 0.029 | 0.4 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vršanská, M.; Voběrková, S.; Jiménez Jiménez, A.M.; Strmiska, V.; Adam, V. Preparation and Optimisation of Cross-Linked Enzyme Aggregates Using Native Isolate White Rot Fungi Trametes versicolor and Fomes fomentarius for the Decolourisation of Synthetic Dyes. Int. J. Environ. Res. Public Health 2018, 15, 23. https://doi.org/10.3390/ijerph15010023
Vršanská M, Voběrková S, Jiménez Jiménez AM, Strmiska V, Adam V. Preparation and Optimisation of Cross-Linked Enzyme Aggregates Using Native Isolate White Rot Fungi Trametes versicolor and Fomes fomentarius for the Decolourisation of Synthetic Dyes. International Journal of Environmental Research and Public Health. 2018; 15(1):23. https://doi.org/10.3390/ijerph15010023
Chicago/Turabian StyleVršanská, Martina, Stanislava Voběrková, Ana María Jiménez Jiménez, Vladislav Strmiska, and Vojtěch Adam. 2018. "Preparation and Optimisation of Cross-Linked Enzyme Aggregates Using Native Isolate White Rot Fungi Trametes versicolor and Fomes fomentarius for the Decolourisation of Synthetic Dyes" International Journal of Environmental Research and Public Health 15, no. 1: 23. https://doi.org/10.3390/ijerph15010023